
Sensitivities and Key Vulnerabilities: Natural Capital
3.1 WATER RESOURCES
Changing climate is, and will continue to be, reflected in the key variables governing the hydrological cycle: temperature, evapotranspiration, precipitation, and snow and ice. The natural health and wealth of the Prairies are intimately linked to the quality and quantity of the water. It impacts almost every aspect of society: health and well-being, agriculture, food production and rural life, cities and infrastructure, energy production and cost, forestry, recreation and other sectors. Some of the greatest societal stresses endured in the Prairies have been directly related to extremes in the hydrological climate system.
Surface Water Resources
Changes in winter precipitation, temperature and duration will have substantial impacts on surface water supplies. Winter warming will reduce snow accumulations in alpine areas (Leung and Ghan, 1999; Lapp et al., 2005) and across the Prairies. This will cause declines in annual streamflow and a notable shift in streamflow timing to earlier in the year, resulting in lower late-season water supplies. Runoff from small prairie and parkland rivers is almost exclusively due to snowmelt runoff over frozen soil (Byrne, 1989). Reduced winter snowfall in the latter half of the twentieth century (Akinremi et al., 1999) contributed to the observed trend of declining streamflows. This is already a critical issue for many rivers in the southern Prairies, such as the Bow, Oldman and Milk, particularly in dry years.
Monthly mean streamflow at 50 gauges in the Prairies showed positive trends in March and negative trends in the autumn months between the late 1940s –early 1950s and 1993 (Gan, 1998). This increase in the March streamflow can be attributed both to an increase in spring precipitation as rain and to earlier snowmelt (Burn, 1994; Gan, 1998; Zhang et al., 2001; Yue et al., 2003). Burn (1994) found that 30% of unregulated rivers in western Canada showed a statistically significant trend towards earlier spring runoff by the 1990s, with greater advances in peak runoff observed in rivers at higher latitudes.
Widespread glacier retreat during the last century has resulted in a measurable decline of summer and fall runoff, impacting rivers during the period of lowest water flow and highest water demand (Demuth and Pietroniro, 2003). Continued glacier retreat will exacerbate water shortages already apparent in many areas of Alberta and Saskatchewan during drought years. Demuth and Pietroniro (2003, p. iv.) concluded that:
“The reliability of water flow from the glaciated headwater basins of the upper North Saskatchewan River Basin has declined since the mid-1900s. Hydrologic and ecological regimes dependent on the timing and magnitude of glacier-derived meltwater may already be experiencing the medium-long-term impacts of climate change discussed by the IPCC.”
Since headwaters of the upper North Saskatchewan River are close to those of the Athabasca River and Bow River systems, all three river systems are likely experiencing the same changes in streamflow now, and will continue to do so in future.
Most scenarios suggest higher winter and spring flows, when more precipitation, especially rainfall, is projected. Reductions in flow are generally projected for the summer, the season of greatest demand for surface water. Pietroniro et al. (2006) coupled hydrological models and climate change scenarios to estimate the following mean annual changes in flow by the 2050s:
- Red Deer River at Bindloss: –13%
- Bow River at the mouth: –10%
- Oldman River at the mouth: –4%
- South Saskatchewan River at Lake Diefenbaker: –8.5%.
Groundwater Resources
Groundwater is the source of potable water for about 21% of Manitoba residents, 23% of Albertans and 43% of Saskatchewan ’s population (Environment Canada, 2004b). Future groundwater supplies will decline in some regions but may increase in others, reflecting a dynamic equilibrium between recharge, discharge and groundwater storage (Maathuis and Thorleifson, 2000; Chen et al., 2002). Increased rainfall in early spring and late fall will enhance recharge if soil water levels are high; otherwise, water will be retained in the soil, benefiting ecosystem and crop productivity. Drier soils due to higher rates of evapotranspiration result in decreased recharge, which would lead to a slow but steady decline in the water table in many regions. When groundwater levels decline due to reduced recharge, prolonged drought or overpumping, water supply and quality are adversely affected. This occurred in central Alberta during the 2001 –2003 drought, which was the most intense long-duration drought in recorded history (Kienzle, 2006). Declines in groundwater levels in a carbonate aquifer near Winnipeg could cause salinization of water wells as the saline-freshwater boundary potentially moves eastward in response to climate change (Chen et al., 2002).
Water Demand
Increases in the demand for water will compound issues of water supply. The magnitude of potential demand is clearly illustrated in the following estimates for the South Saskatchewan River basin (Alberta Environment, 2002, 2005, 2006):
- Demand from non-irrigation could increase between 35% and 67% by 2021 and between 52% and 136% by 2046.
- Irrigation districts have the potential to expand by up to 10% and 20% in the Oldman River and Bow River basins, respectively.
- Population in the South Saskatchewan River basin is expected to grow from 1.3 million in 1996 to more than 2 million by 2021, and to more than 3 million by 2046.
- About 65% of the average natural flow of the Oldman River and its tributaries has been allocated (Government of Alberta, 2006). Most of these allocations (almost 90%) are committed to irrigated agriculture. New water license allocations are not available in the Oldman River and Bow River basins, so all growth must occur within existing allocations.
Water Quality
Aquatic ecosystems and water resources in the Prairies face a range of threats related to water quality that will be exacerbated by climate change. These include (Environment Canada, 2001):
- physical disruptions associated with
- land-use impacts of agriculture and forestry,
- urban water withdrawals
- sewage effluent and storm water runoff, and
- impacts of dams and diversions;
- chemical contamination, including
- persistent organic pollutants and mercury,
- endocrine-disrupting substances,
- nutrients (nitrogen and phosphorus),
- urban runoff and municipal wastewater effluent, and
- aquatic acidification; and
- biological contamination, such as waterborne pathogens.
Reductions in streamflow under climate warming will worsen these impacts. Dilution capacity will likely decline as streamflows decrease and lake residence times increase accordingly. Droughts could result in enhanced soil erosion from both agricultural lands and areas burned by forest fires. This erosion will increase stream sediment loads and enhance nutrients in local water systems, leading to eutrophication of water bodies and increased pathogen loading in streams during the summer (Hyland et al., 2003; Johnson et al., 2003; Little et al., 2003). The Millennium Ecosystem Assessment (2005) identified the joint effects of climate change and nutrient overenrichment as the major threat to drylands agro-ecosystems. The size of the massive algae blooms in Lake Winnipeg correlate with higher summer temperatures (McCullough et al., 2006). Since the Lake Winnipeg watershed encompasses much of the Prairie Ecozone, the lake receives runoff from a large proportion of Canada ’s agricultural land.
Impacts of Extreme Hydrological Events
The few studies that have examined GCM outputs for extremes of future climate (e.g. Kharin and Zwiers, 2000) suggest an increased probability of extreme conditions, including a greater frequency of flooding and severe drought. A likelihood of increased drought severity is also inferred from both recent and prehistoric conditions of the western interior. The recent climate (since the 1940s) has been characterized by severe droughts (large water deficits) of relatively short duration compared to preceding centuries (Sauchyn et al., 2003). The natural climate cycles that underlie climate change include drought of much longer duration than those that occurred in the twentieth century (see Section 2.4).
In the boreal forest and taiga areas, increased drought frequency, including persistent multi-year droughts (Sauchyn et al., 2003), will result in declining soil water and increased forest fire extent and net area burned. During recent extreme droughts, organic soils have dried and burned with forests, resulting in an almost total loss of vegetation and soil cover, and subsequently the ability to store water locally. Under these conditions, runoff becomes instantaneous, resulting in flash floods.
Increasing annual and seasonal temperatures will exacerbate drought conditions (Laprise et al., 2003), but warmer temperatures also increase the likelihood of extreme rainfall events (Groisman et al., 2005). Such events frequently cause local or regional flooding, such as that experienced in the South Saskatchewan River basin in 1995 and 2004. These and the other recent flood and drought events listed below demonstrate the pressures on water resource management that are expected to increase in future:
- An Edmonton thunderstorm dropped 150 mm of rain in under an hour on a city already saturated by earlier storms. Losses were estimated at $175 million (Environment Canada, 2004a).
- The instrumental records indicate that the recent sustained drought of 2000–2003 recorded far less total precipitation than that of the mid-1930s (Kienzle, 2006). The most severe impacts on soil moisture and groundwater levels are from multi-year droughts.
- Environment Canada (2004a) characterized the 2002 crop year as “the worst ever for farmers in Western Canada.”
- In 2001, the St. Mary River Irrigation Project in southern Alberta had insufficient water to meet annual allocations: farms were only provided with 60% of their water allocations.
Adaptation
Adaptation to changes in the hydrology of the Prairies will be challenging, especially where current water supplies are almost fully allocated. Future water scarcity could lead to abandonment and/or underutilization of major infrastructure (canals, pipelines, dams and reservoirs) worth billions of dollars. Rising demand due to warmer climate and a decline in summer runoff in some years will lead to calls for increased storage and diversion of water away from areas with water surpluses. However, reservoirs are greenhouse gas sources (St. Louis et al., 2000), and dams and diversions have well-documented negative environmental impacts (Environment Canada, 2001; Mailman et al., 2006). Adaptation in the water resources sector is also discussed in Section 5.1.1.
3.2 ECOSYSTEMS
In a global analysis, climate change is rated as second only to land use in importance as a factor that is expected to determine changes in biodiversity during the current century (Sala et al., 2000). Changes in climate will alter environmental conditions to the benefit of some species and the detriment of others, often with economic consequences. For example, as vegetation and insects shift in response to changing climate, tourism and recreation activities such as bird watching will be affected, and agricultural, forestry and urban pest management practices may have to adjust.
Biodiversity and Productivity
In the absence of moisture limitations or other constraints, plant productivity should rise with an increase in temperature and length of growing season. Increased photosynthetic activity for much of Canada during the period 1981 –1991 has been attributed to a longer growing season (Myneni et al., 1997). Little is known, however, about which species or assemblages will be relatively advantaged or disadvantaged in increasingly moisture-constrained ecosystems. Changes in the timing and intensity of freeze-thaw events, diurnal temperature patterns (Gitay et al., 2001), and storm and wind stress events may influence vegetation distribution or survival, especially of various tree species (Macdonald et al., 1998), but the details of how this will occur are not known.
Factors other than temperature and precipitation will also affect prairie ecosystems. For example, CO 2 fertilization increases the efficiency of water use by some plant species (Lemon, 1983), although there are many uncertainties about the sum effect (Wheaton, 1997). While studies have reported a positive CO 2 enrichment effect on the growth of white spruce in southwestern Manitoba (Wang et al., 2006), modelling and an empirical study (Gracia et al., 2001) suggested that any positive CO 2 fertilization effect is neutralized among evergreens because growth is constrained by moisture limitations. Prediction of overall changes in forest CO 2 uptake and storage, independent of interspecies variations, is not yet possible (Gitay et al., 2001). One major problem in predicting the impacts of CO 2 enrichment on a specific species is that the impact occurs on all vegetation simultaneously. It is not enough to know the CO 2 response of one species; rather, one needs to know the relative growth advantage, if any, gained by all vegetation species competing for resources at a given site.
Ultraviolet B radiation and ground-level ozone levels are increasing and are expected to negatively impact vegetation, possibly nullifying any positive CO 2 enrichment effect (Henderson et al., 2002). In addition, nitrogen deposition from industrial activity may be affecting species growth and competitive interactions, even in locations far from industrial centres (Kochy and Wilson, 2001).
Changes in forest disturbance regimes induced by climate change may be substantial enough to alter current forest ecosystems (Loehle and LeBlanc, 1996). Henderson et al. (2002) noted two pathways of forest change: 1) slow and cumulative decline; or 2) catastrophic loss, such as a major fire. Increased average winter temperatures will lead to greater overwinter survival of pathogens and increased disease severity (Harvell et al., 2002). Drought conditions weaken trees ’ defences to more virulent pathogens (Saporta et al., 1998). As conditions become more xeric, the lifespan of conifer needles is reduced, placing conifers under increasing stress (Gracia et al., 2002). The boreal forest is expected to be significantly affected by climate change, especially at its southern boundary (Herrington et al., 1997; Henderson et al., 2002; Carr et al., 2004). Major changes in species representation are projected for Saskatchewan ’s boreal forest by 2080 through impact modelling (utilizing the CGCM1 and the A1 emission scenario; Carr et al., 2004).
Changes in the Timing of Biological Events
Early settlers and Aboriginal people recognized the timing of biological events as a function of season and weather, and used these indicators to forecast the timing and success of planting, fishing and hunting activities (Lantz and Turner, 2004). The dates and rates of spring flowering of widely distributed wild plants are among the most reliable events that can be monitored and used as an index of weather and climate. A program called ‘Plantwatch’ monitors the phenology of flowering of key wild plants through the reports of a network of volunteers, and has become an important tool for tracking the impacts of changing climate (Beaubien, 1997). Dates of flowering of key perennial plants in Alberta are closely related to the average temperature two months prior to bloom (Beaubien and Freeland, 2000). A 26-day shift to earlier onset of spring has already occurred over the past century (Beaubien and Freeland, 2000). The spring flowering index derived from the Plantwatch data was found to be correlated with Pacific sea-surface temperatures, including El Ni ño events.
Vegetation Zone Response
Models of vegetation zonation have shown a northward shift of the forest-grassland boundary in the Prairies with climate change (Hogg and Hurdle, 1995; Vandall et al., 2006). Grasslands are also projected to change, with aspen parkland and fescue prairie of the present northern fringe giving way to variants of mixed prairie. These modelled shifts in zonation do not specify the exact composition of future vegetation because of lags in migration of some species. However, the following trends are projected from the present to the 2050s (Vandall et al., 2006):
- In forest regions, there will be a general reduction in tree growth, regeneration failure in dry years and a gradual reduction in tree cover and expansion of grassland patches.
- In the aspen parkland, there will be shrinking of aspen groves, reduced invasion of grassland patches by shrubs and poplar sprouts, and decreasing shrub cover.
The most significant impacts can be expected to occur at the interfaces of drier grassland with the moister foothills grassland, and at the interface of grassland with parkland and forest.
Grassland production is limited by moisture supply. Although the warmer and drier climate projected for the Prairies would suggest declining production and grazing capacity, actual changes in grassland production are likely to be modest, given a longer growing season, reduced competition from shrubs and trees, and increases in warm-season grasses that have higher water-use efficiency (Thorpe et al., 2004).
Impact models that consider simply the current (static) positions of ecoregions (e.g. Davis and Zabinski, 1992) have projected significant changes in boreal forest area and quality. Models based on plant growth and population dynamics would yield more robust predictions. The northern extremes of the boreal forest will likely extend under climate warming, but the rate of northern extension of the forest is uncertain, and will take decades as trees respond to variations in soil temperature, permafrost and uncertain seed dispersal and establishment (Lloyd, 2005). An associated change in the southern boundary of the forest is likely, but would be influenced by droughts and associated large-scale fire events.
Estimating the timing of ecological changes is difficult, in part because of inability to predict precise thresholds. Vegetation responds after the fact to climate change (autonomous adaptation), and it is natural for a given ecosystem to be ‘behind’ environmental conditions to some degree, a condition termed ecological inertia (see Henderson et al., 2002). Anderson et al. (1997) warned that ecosystems can absorb stresses over long periods of time before crossing a critical threshold, which may lead to rapid ecosystem and landscape modification. The climate change impact on mature trees is not likely to be noticeable until biological thresholds are reached and dieback results (Saporta et al., 1998).
Climate change will be significant in all of the prairie-parkland national parks (Elk Island, AB; Prince Albert, SK; Riding Mountain, MB) and Wood Buffalo National Park, NT (Scott and Suffling, 2000). These parks can expect increases in forest fire frequency and intensity, increased forest disease outbreaks and insect infestations, and loss of boreal forest to grassland and temperate forest (de Groot et al., 2002). Climate change represents “an unprecedented challenge for Parks Canada” and “current ecological communities will begin to disassemble and ‘resort’ into new assemblages” (Scott and Suffling, 2000). To address this challenge, Henderson et al. (2002, p. 3) stated that “In a world of climate change, selection of protected areas may need to focus on site heterogeneity and habitat diversity (as these provide some buffer against climate change) rather than on representativeness. ” For example, high-relief terrain, such as the Cypress Hills landscape, can always be expected to provide a range of habitats and ecosystems different from the surrounding plains, and therefore contribute to biodiversity, even as the nature of these habitats and ecosystems changes over time (see Case Study 1). However, a low-relief landscape, such as Prince Albert National Park, which is mandated to protect fescue grassland, aspen parkland and southern boreal forest within the national parks system, may fail to preserve these landscape elements over time, just as Wapusk National Park, MB may fail to protect polar bear habitat, its mandated raison d'être.
Wildlife
The prairie pothole region of central North America is the single most productive habitat for waterfowl in the world, with the Canadian Prairies providing breeding grounds for 50 to 80% of the Canadian duck population (Clair et al., 1998). Increasing aridity in the prairie grasslands is likely to negatively impact migratory waterfowl populations (Poiani and Johnson, 1993) as waterfowl numbers decrease in response to drought and habitat loss (Bethke and Nudds, 1995). Weather fluctuations during the breeding season account for more than 80% of the variation in population growth rate of mallards and other ducks (Hoekman et al., 2002). In northern regions, earlier dates of disappearance of snow and increasing average temperatures have resulted in earlier nesting and hatching of geese (e.g. LaRoe and Rusch, 1995).
Wildlife migration patterns and population size have already been affected by recent climate trends, and further impacts are expected with climate change (Inkley et al., 2004). This will affect hunting-based industries, environmental activities, fishing regulations and production, and traditional ways of life reliant on vertebrate biodiversity. Relatively sessile animals, such as reptiles and amphibians, are at greater risk of extirpation than relatively mobile birds or butterflies. Aquatic ecosystems will be stressed by warmer and drier conditions, and a large number of prairie aquatic species are at risk of extirpation (James et al., 2001). Many fish species, for example, are sensitive to small changes in temperature, turbidity, salinity or oxygen regimes. For the Prairies, larger algal blooms, accelerated eutrophication and serious impacts on fish species are expected, due to a combination of climate change, increasing nutrient runoff and increasing human use pressures on natural water systems (Schindler and Donahue, 2006).
Adaptation
Conservation policy can aim to extend ecological inertia, have no impact on it or reduce it (Henderson et al., 2002). Vegetation associations that are most ‘in tune’ with the evolving climate will require the least degree of human intervention. Conversely, those vegetation ensembles that are outside their natural climate norms will require increasingly intensive and active human intervention and management to survive. However, with a high degree of human intervention, it will be possible at some sites to maintain vegetation (and associated fauna) that would otherwise certainly disappear.
CASE STUDY 1
Climate Change Impacts on the Island Forests of the Great Plains

Larger image
[JPEG, 518 X 168, ]
{kind=link}
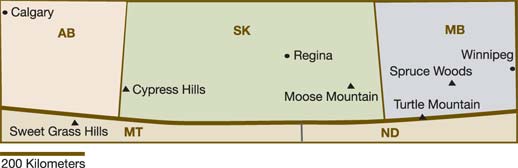
Scattered from the plains of central Alberta to Texas are island forests: refugia of trees and tree-dependent species isolated in a sea of grass. Henderson et al. (2002) examined the impacts that climate change will have on five of these ecosystems in the southern Prairies and adjacent North Dakota and Montana: the Cypress Hills, Spruce Woods, Turtle Mountain, Moose Mountain and the Sweet Grass Hills (Figure 11). These island forests have considerable regional significance in terms of biodiversity, wildlife habitat, grazing land and sources of timber, and as the headwaters of many prairie streams.
Plains island forests are at risk from climate change. They are marginal or ecotone systems, borderline between grassland and forest ecosystems, and therefore sensitive to small changes in environmental conditions. As they are relatively small ecosystems, island forests may exhibit lower genetic diversity and greater vulnerability to catastrophic disturbance, such as wildfire, pathogen attack or severe drought.
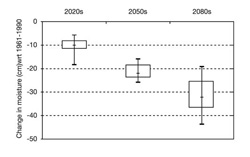
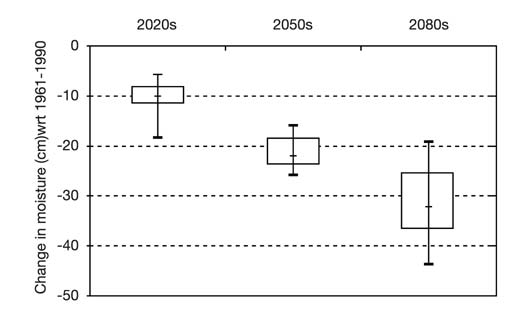
FIGURE 12: Summary of the projected changes in soil moisture levels (averaged over the five island forest study sites) for the 2020s, 2050s and 2080s. The thin vertical lines in the plot indicate the range of possible future moisture levels compared with the climate of 1961-1990. The boxes indicate the moisture level ranges within which 50% of the scenario projections fall. The horizontal dash within each box indicates the median moisture scenario.
Larger image
[JPEG, 531 X 334, ]
{kind=link}
The study used a range of climate scenarios derived from three global climate models (HadCM3, CGCM2, and CSIROMk2b) to determine the future moisture regimes for the five island forests and to consider the implications of these moisture regimes for the dominant tree species. In the plains, a region always on the edge of drought stress, soil moisture levels represent the single most important climate change parameter for natural ecosystems. The net effect on moisture levels of the modeled changes in both temperature and precipitation is shown in Figure 12. Increased temperatures will have a powerful evaporation effect, such that soil moisture balances will decline substantially.
Henderson et al. (2002) concluded that the island forests will suffer serious challenges to ecosystem integrity. Highly intensive management will likely be necessary to preserve some type of forest cover at these sites. Management that aims simply to retain existing vegetation, or to restore historical vegetation distributions and ecosystems, will fail as the climate moves farther away from recent and current norms. Possible adaptation actions range from protecting the forests by maintaining a diversity of age stands and responding aggressively to pathogen disturbances, to actively regenerating the forest with existing or alien tree species that are better adapted to the new climate parameters.
Increasing connectivity between protected areas to facilitate migration of particular species populations is commonly proposed as one method of coping with climate change (e.g. Malcolm and Markham, 2000; James et al., 2001; Joyce et al., 2001). Although some species might be able to migrate, others will be threatened by the arrival of new competitors or by the pathogens that increased connectivity supports. Thus, increased connectivity may also hasten the decline of some ecosystems by favouring alien invasions.
Intentional introduction of a species native to a nearby area may be a useful tool for adaptation to climate change, and such introductions are less likely to be ecologically disruptive than Eurasian exotic species (Thorpe et al., 2001). Yet the most drought-hardy trees that could potentially survive in the northern plains derive from central Asia (Henderson et al., 2002). Western conifers, such as Douglas fir and ponderosa pine, and hardwoods of the southern Prairies, such as Manitoba maple and green ash, may be suited to future climates of the western Boreal Ecozone (Thorpe et al., 2006). Native boreal species, on the other hand, are expected to shift northward and decline in the southern parts of their current range.
In restoration ecology, “it is often assumed that understorey vegetation will establish over time (‘plant trees and the rest will come’), but natural invasion may not automatically bring back all species desired” (Frelich and Puettmann, 1999). However, little is known about alien tree introductions in prairie forests, and virtually nothing about mid- and understorey introductions. Biodiversity managers need to think of themselves not as practitioners of preservation, but as ‘creation ecologists’, as antecedent landscapes can no longer be effectively targeted. We have options, but the past is not one of them. The future ecosystems that result from climate change in the Prairies will be unprecedented.
3.3 SOIL LANDSCAPES
The Rocky Mountains of western Alberta and the dryland landscapes of the Prairie Ecozone are highly dynamic and active landscapes. Catastrophic and hazardous geological processes associated with extreme climate events are common on the long, steep slopes of the Canadian Cordillera. For example, in August 1999, a debris flow at Five-Mile Creek in Banff National Park blocked the Trans-Canada Highway for several days at the peak of the tourist season (Evans, 2002). Such events are nearly always triggered by excess rainfall or by runoff from rapid melting of snow or ice. An increased frequency of landslides, debris flows, rock avalanches and outburst floods is probable, given current and projected future trends in hydrology and climate that include increased rainfall (especially in winter), rapid snowmelt and shrinking glaciers (Evans and Clague, 1994, 1997). These changes will affect public safety and the maintenance of infrastructure, especially with increasing recreational activity and residential development in the Rocky Mountains. In the longer term, further warming, drought and the complete wastage of glaciers could cause catastrophic events to taper off; however, the decay of permafrost could accelerate slope failures at high elevations for many decades (Evans and Clague, 1997).
Most of the Prairies region is underlain by poorly consolidated sediments that erode and fail where they are exposed to the forces of wind, water and gravity on valley sides, and where farming or aridity limit the vegetation cover. The most active landscapes are the dune fields and river valleys (Lemmen et al., 1998) that are sensitive to hydrological and climatic variation and extremes (Lemmen and Vance, 1999). Projected increases in drought and aridity will likely result in more widespread wind erosion and increased sand dune activity (Wolfe and Nickling, 1997). Current dune activity in the dry core of the mixed grassland ecoregion serves as an analogue of the potential response of currently stable dune fields on the moister margins of the Prairie Ecozone and the southern boreal plains (Wolfe and Nickling, 1997). Global climate model –based assessments of future vegetation and soil moisture (Thorpe et al., 2001) suggest that vegetation will shift towards more open grassland, with increased potential for sand dune activity. Climate at the driest sites may exceed thresholds for active sand dune crests, and would require proactive land-use management and stringent enforcement of current guidelines and regulations to limit dune activity. Slopes and stream channels exposed to less frequent but more intense rainfall will also be subjected to enhanced erosion and shallow slope failures because the protective vegetation cover will suffer from the prolonged dry spells (Sauchyn, 1998; Ashmore and Church, 2001).
With the modification of about 90% of the Prairie Ecozone for agriculture, tens of millions of hectares were exposed to soil erosion. Annual soil loss from cropland is two to three orders of magnitude higher than on rangeland (Coote, 1983). Wind and water erosion are episodic: centimetres of topsoil can be removed during a single event, reversing centuries or millennia of soil formation and seriously reducing the natural fertility of cropland. The semiarid to subhumid mixed grassland ecoregion, an area of approximately 200 000 km 2, is at risk of desertification. The human impacts on prairie soils are reduced by soil conservation but can be exacerbated by social and economic factors, including the declining national significance of prairie agriculture since the 1960s, the rising influence of global market forces and multinational corporations, declining rural population, and reduced support for agricultural research, farm crisis relief, income support programs and grain transportation (Knutilla, 2003).
Whereas social and economic futures are not easily predicted (see Section 2.3), projections of future aridity can be derived from GCMs. When the Aridity Index (ratio of precipitation to potential evapotranspiration, or P/PET) was computed for 1961 to 1990 and for the 2050s, using output from the Canadian CGCM2 and emission scenario B2, the area of land at risk of desertification (P/PET <0.65; Middleton and Thomas, 1992) increased by about 50% (Sauchyn et al., 2005). In the scatterplots of future climate (Figure 8), this scenario would plot above the median temperature and below the median precipitation; it is a moderately warm and dry climate scenario. With the high inter-annual variability in prairie climate, the trend towards increased aridity will be realized with droughts that are more frequent and/or sustained than the intervening years of normal to above-average moisture. Prolonged droughts are likely to exceed soil moisture thresholds beyond which landscapes are more vulnerable to disturbance and potential desertification (Prairie Farm Rehabilitation Administration, 2000).
Page details
- Date modified: